5 Mamíferos terrestres de médio e grande porte e aves terrícolas cinegéticas
Arlindo Gomes Filho1, Elildo Alves Ribeiro de Carvalho Junior2, Gerson Buss3, Marcelo Lima Reis4, Marcos de Sousa Fialho1, Rafael Suertegaray Rossato3, Ricardo Sampaio3, Richard Hatakeyama5 & Thiago Orsi Laranjeiras6
Centro Nacional de Pesquisa e Conservação de Aves Silvestres – CEMAVE
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
BR-230 Km 10
Floresta Nacional da Restinga de Cabedelo
58108-012 Cabedelo, PBCentro Nacional de Pesquisa e Conservação de Mamíferos Carnívoros – CENAP
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
Estrada Municipal Hisaichi Takebayashi, 8600 - Bairro da Usina
12952-011 Atibaia, SPCentro Nacional de Pesquisa e Conservação de Primatas Brasileiros – CPB
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
BR-230 Km 10
Floresta Nacional da Restinga de Cabedelo
58108-012 Cabedelo, PBCoordenação de Monitoramento da Biodiversidade - COMOB
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
Complexo Administrativo EQSW 103/104 s/n
70670-350 Brasília, DFNúcleo de Gestão Integrada ICMBio Tefé
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
Estr. do Aeroporto, 725 - Centro
69550-101 Tefé, AMNúcleo de Gestão Integrada ICMBio Roraima
Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio
Rua Alfredo Cruz, 283 - Centro 69301-140 Boa Vista, RR
Como citar: Gomes-Filho, A.; Carvalho-Jr., E. A. R.; Buss, G.; Reis, M. L.; Fialho, J. M. S.; Rossato, R. S.; Sampaio, R.; Hatakeyama, R. & Laranjeiras, T. O. Mamíferos terrestres de médio e grande porte e aves terrícolas cinegéticas. In: Programa Nacional de Monitoramento da Biodiversidade – Programa Monitora, Subprograma Terrestre, Componente Florestal. Relatório 2014-2022. Instituto Chico Mendes de Conservação da Biodiversidade, Brasília, 2024.
Mamíferos terrestres de médio e grande porte e aves terrícolas cinegéticas são animais sensíveis à intensa pressão de caça, redução e fragmentação de habitat, e por isso são considerados bons indicadores de impactos de origem antrópica. A defaunação das florestas acarreta a chamada “síndrome da floresta vazia”, a ausência de muitas espécies de animais causada pela forte pressão humana e seus efeitos (Redford 1992).
A redução das populações animais afeta diretamente a biodiversidade e impacta negativamente a própria estrutura e sobrevivência das florestas, em razão da perda de polinizadores, dispersores de sementes e outros processos ecológicos essenciais. Populações de grandes herbívoros, por exemplo, apesar de promoverem e regularem a diversidade de plantas e a ciclagem de nutrientes, na ausência de predadores podem crescer de forma descontrolada e gerar uma paisagem de intensa herbivoria, com aumento da supressão vegetal, da predação de sementes e da mortalidade de plântulas, o que reduz a taxa de regeneração da vegetação (Pires & Galetti 2023).
O desequilíbrio na diversidade de espécies em direção a ambientes mais simples, além da redução da biodiversidade em si, pode também contribuir para surtos de doenças e até mesmo a ocorrência de futuras pandemias (Morse et al. 2012). Nesse contexto, o monitoramento desses dois grupos animais em programas como o Monitora é de fundamental importância para a saúde do meio ambiente e das pessoas.
Nesse capítulo apresentamos alguns resultados gerais do monitoramento de mamíferos terrestres de médio e grande porte e aves terrícolas cinegéticas, sistematizados até o momento pelo Programa Monitora. A taxonomia utilizada foi a mesma adotada no processo de avaliação do risco de extinção das espécies da fauna brasileira (Instituto Chico Mendes de Conservação da Biodiversidade - ICMBio 2024).

5.1 Implementação
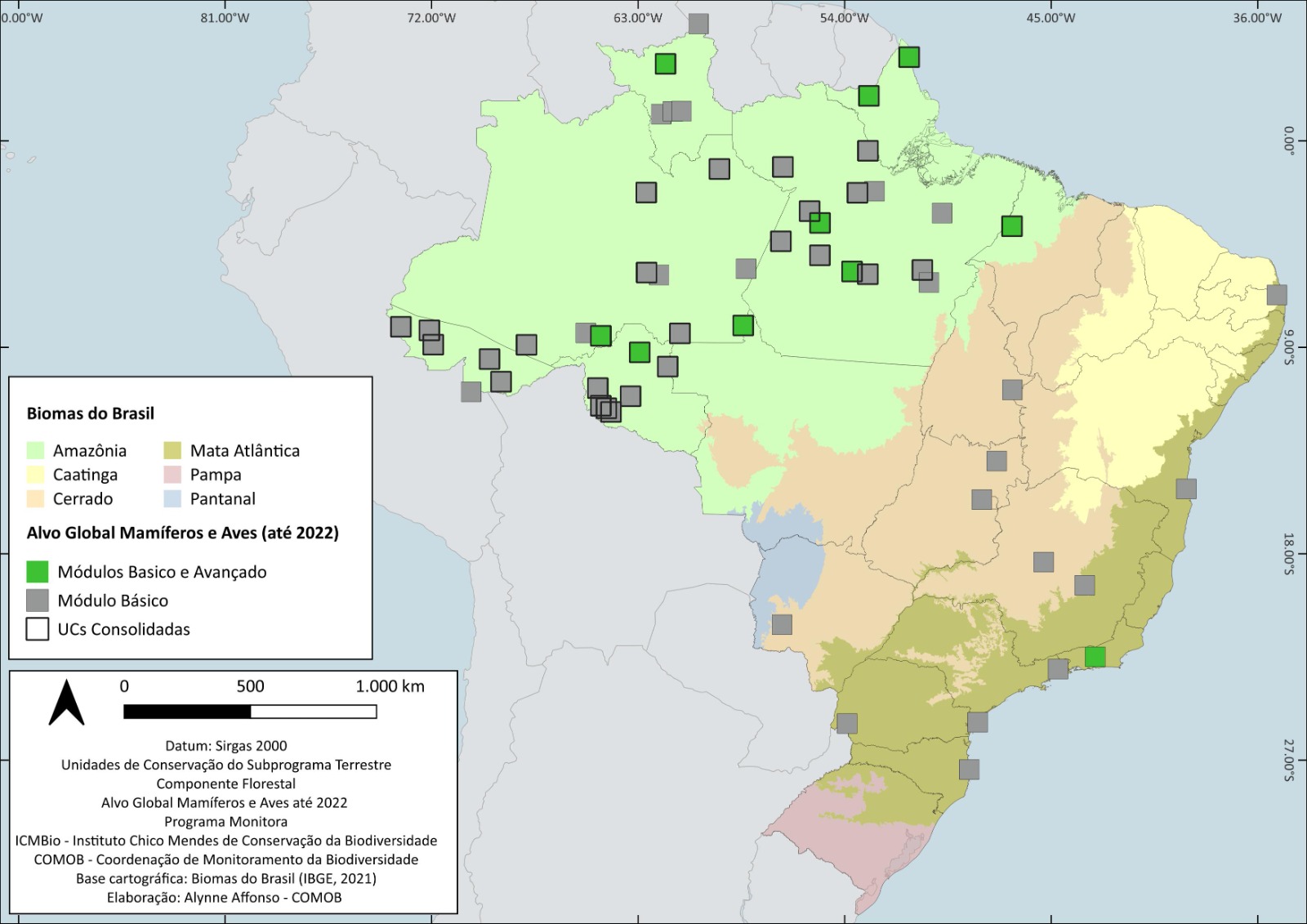
De 2014 a 2022 o protocolo básico de amostragem de mamíferos e aves (censo diurno em transecção linear) foi aplicado em 53 UCs federais1, distribuídas nos biomas Amazônia, Cerrado e Mata Atlântica, totalizando 140 unidades amostrais, que neste protocolo também correspondem às estaçõs amostrais (EAs). Das unidades de conservação amostradas, 33 (63%) já estão consolidadas, com pelo menos três estações amostrais em operação (Figura 5.1). Seis unidades de conservação (12%) retomaram as amostragens em 2022, sete UCs (13%) estão com atividades do Monitora interrompidas (há mais de dois anos consecutivos sem amostragem) e duas (4%) não coletaram dados em 2022 (estão inoperantes).
O esforço de amostragem nos nove anos considerados neste relatório correspondeu a 5.356 dias de campo (transecção/dia) e 25.602,55 km percorridos (Figura 5.2), resultando em 22.985 registros de mamíferos de médio e grande porte e em 12.995 de aves terrícolas cinegéticas.
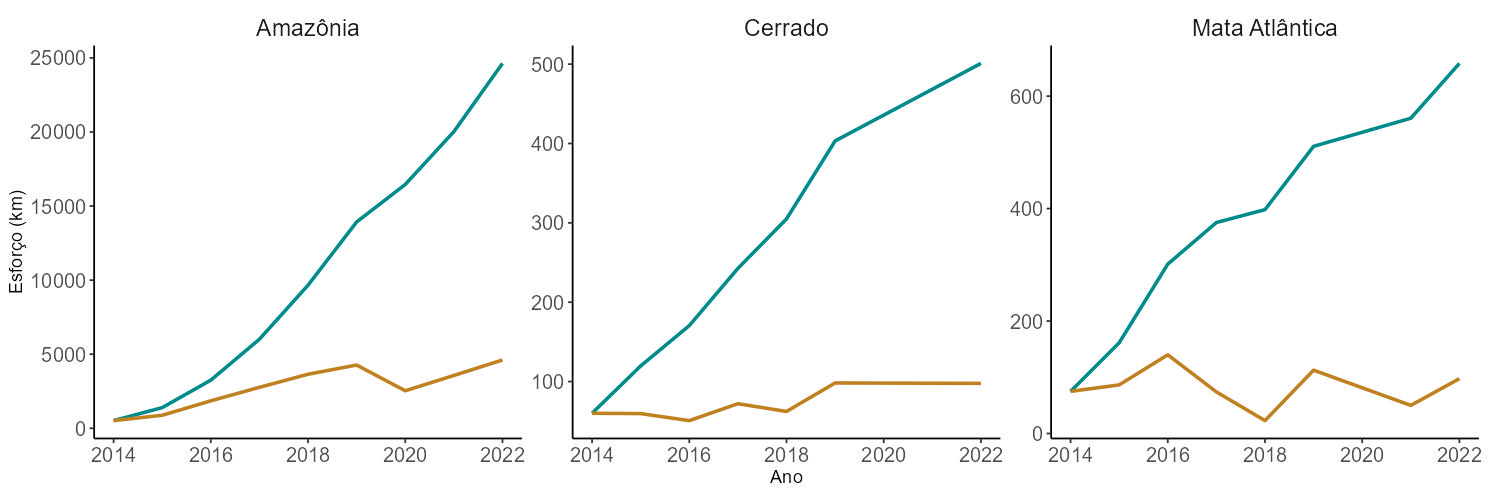
O esforço por UC e por bioma é apresentado na Figura 5.2 e na Figura 5.3. Mais de 95% do esforço de amostragem ocorreu no bioma Amazônico. Isso se deve tanto ao maior número de unidades de conservação integrando o Programa Monitora nesse bioma, quanto ao esforço médio empregado em cada unidade de conservação. As unidades de conservação no Cerrado e na Mata Atlântica são menores e nem sempre comportam a rede recomendada de transecções: três estações amostrais de cinco quilômetros cada. Ademais, custear as atividades de campo nesses biomas é geralmente mais difícil, dada a maior disponibilidade de financiamento internacional para programas de monitoramento na Amazônia.
5.2 Resultados
5.2.1 Visão geral
A maioria dos registros (56%) do protocolo básico de monitoramento de mamíferos e aves correspondeu a primatas e roedores (Dasyproctidae e Sciuridae) (Figura 5.5). Esse resultado se deve, em parte, ao fato de o método ser mais eficiente na detecção desses grupos, que são predominantemente diurnos. Espécies noturnas e esquivas, como a maior parte dos carnívoros, são pouco registradas.
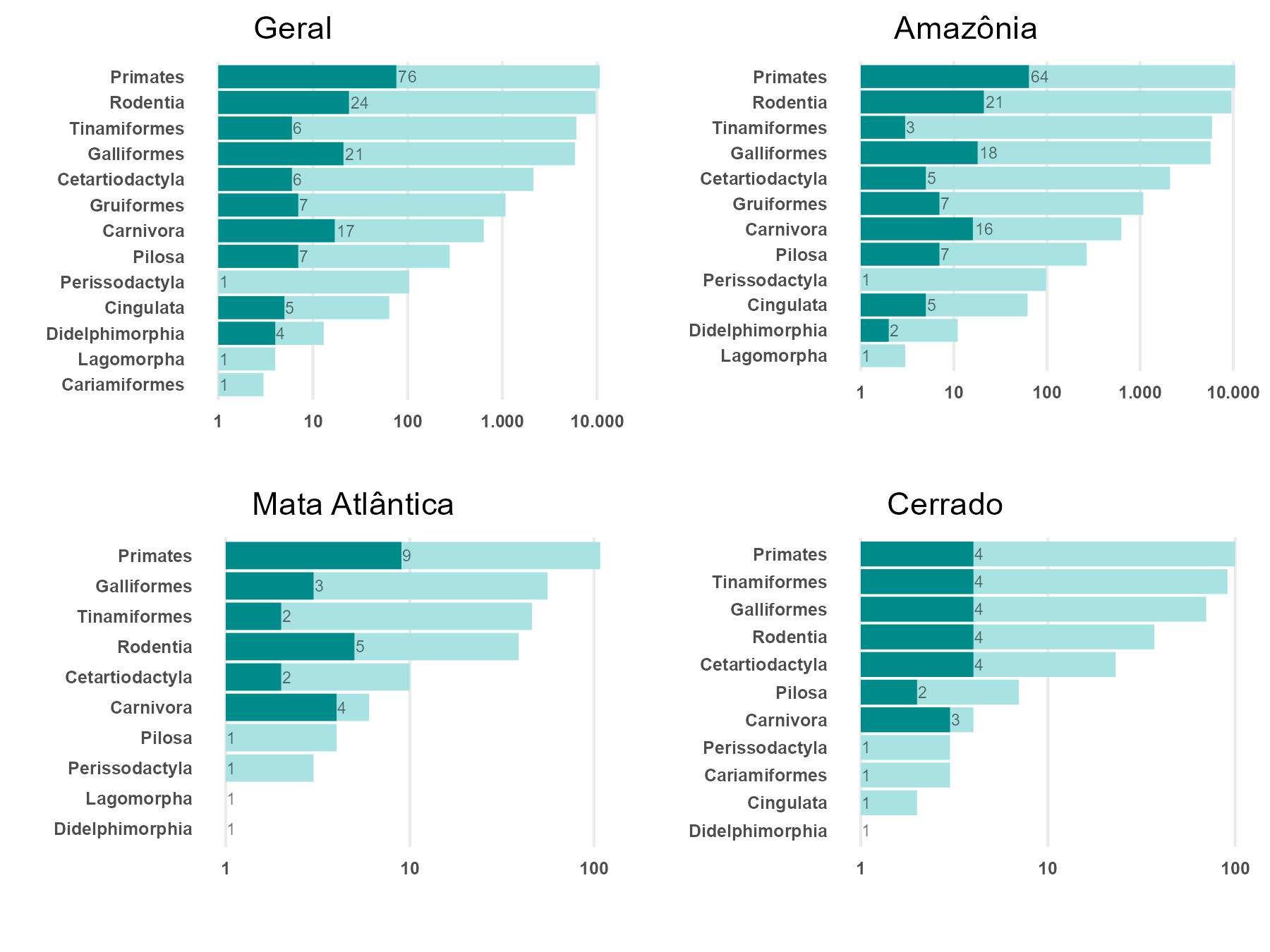
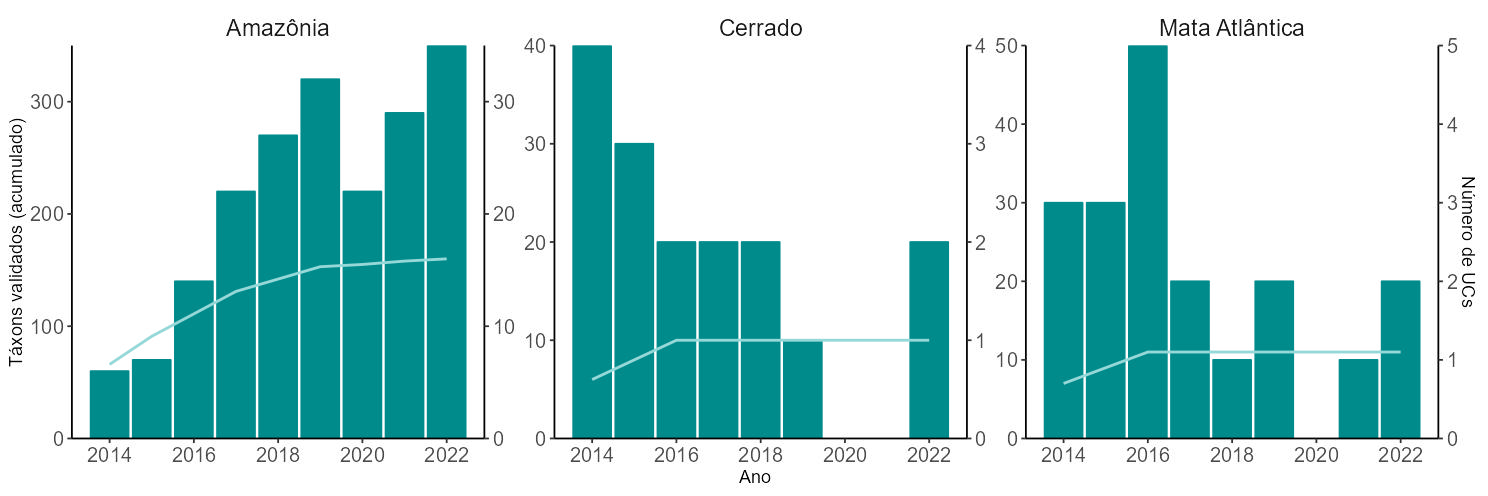
Dentre os mamíferos, os primatas destacaram-se como o grupo preponderante em número de registros e, exceto pelo gênero Callibella (sagui-anão), todos os demais gêneros de primatas com ocorrência no Brasil foram registrados. Setenta e duas espécies foram detectadas, sendo que 18 espécies são consideradas ameaçadas de extinção (Apêndice H). As espécies Callicebus vieirai (Plecturocebus vieirai) e Pithecia irrorata, que eram classificadas como deficientes de dados (DD), passaram a ser categorizadas como “menos preocupante” (LC) no ciclo avaliativo de 2019 (Instituto Chico Mendes de Conservação da Biodiversidade - ICMBio 2024).
Com relação às aves, 29 espécies de 13 gêneros foram registradas (Apêndice I). Durante as amostragens busca-se a identificação dos indivíduos observados em nível específico. Contudo, em algumas unidades de conservação duas, três ou mais espécies muito semelhantes de um mesmo gênero ocorrem em simpatria. Nessas situações, por segurança, esses registros são validados taxonomicamente em nível de gênero. Como exemplos, podemos citar Nothura (codornas), Penelope (jacus), Tinamus (macucos) e Crypturellus (inhambus), todos gêneros com um ou mais táxons ameaçados de extinção (Apêndice I), conforme a Portaria MMA nº 148/2022.
A maioria dos registros de aves distribui-se quase que igualmente entre as ordens Galliformes e Tinamiformes. Esse resultado se deve ao fato de o método ser eficiente na detecção desses grupos e em razão dos Gruiformes (jacamins) ocorrerem apenas no bioma Amazônico, nunca com mais de uma espécie por localidade, enquanto os Cariamiformes, com uma única espécie (Cariama cristata - seriema), são típicos de ambientes abertos e praticamente não foram avistados (Figura 5.5).
A variação nas proporções de registros no Cerrado e na Mata Atlântica ao longo dos nove anos de amostragem deve-se ao ainda pequeno número de unidades de conservação desses biomas integrando o Programa Monitora, à inconstância nas amostragens e ao pequeno esforço amostral, seja por unidade de conservação, seja por bioma (Figura 5.5).
5.2.2 Abundância relativa de mamíferos e aves por biomas
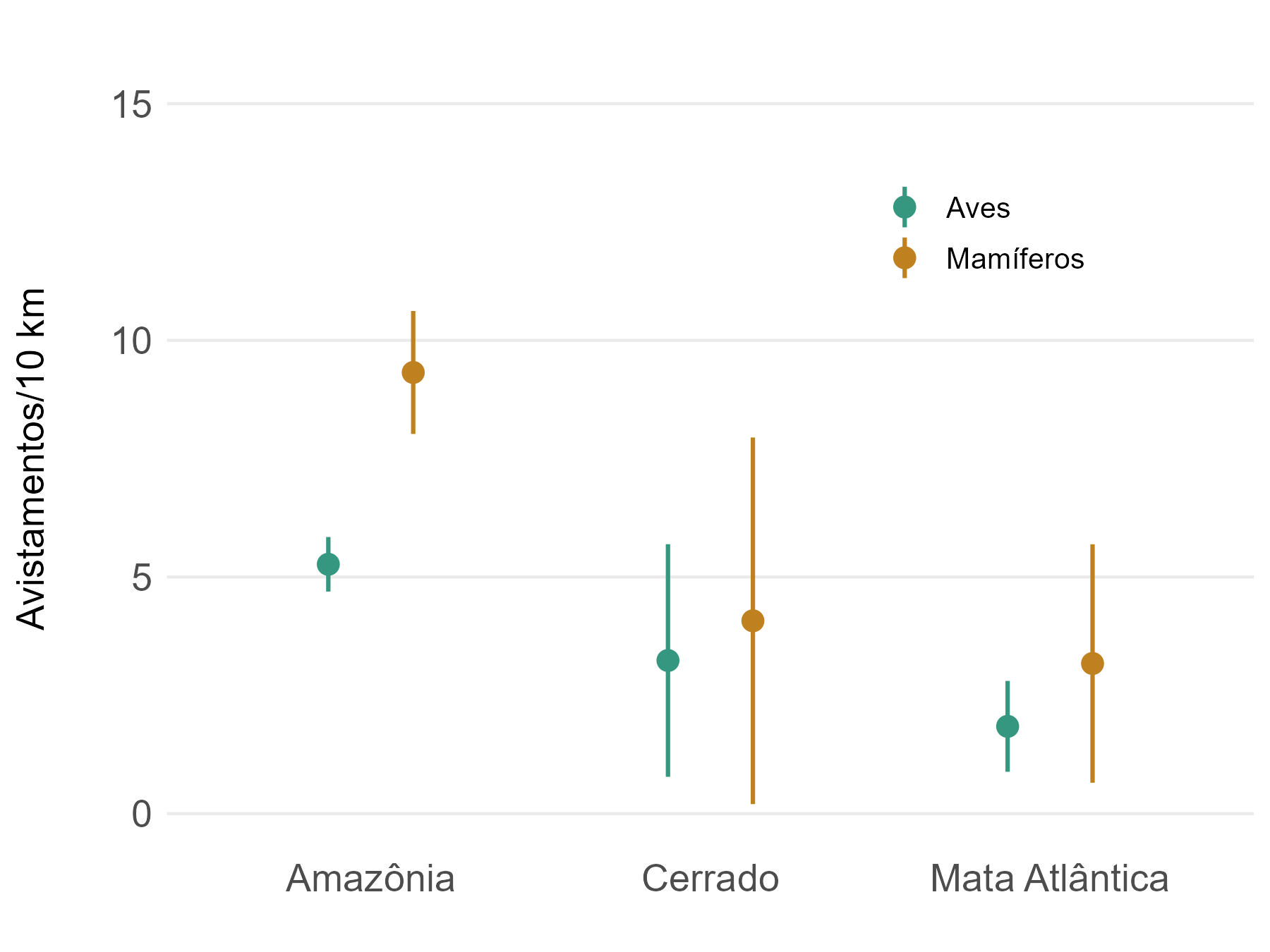
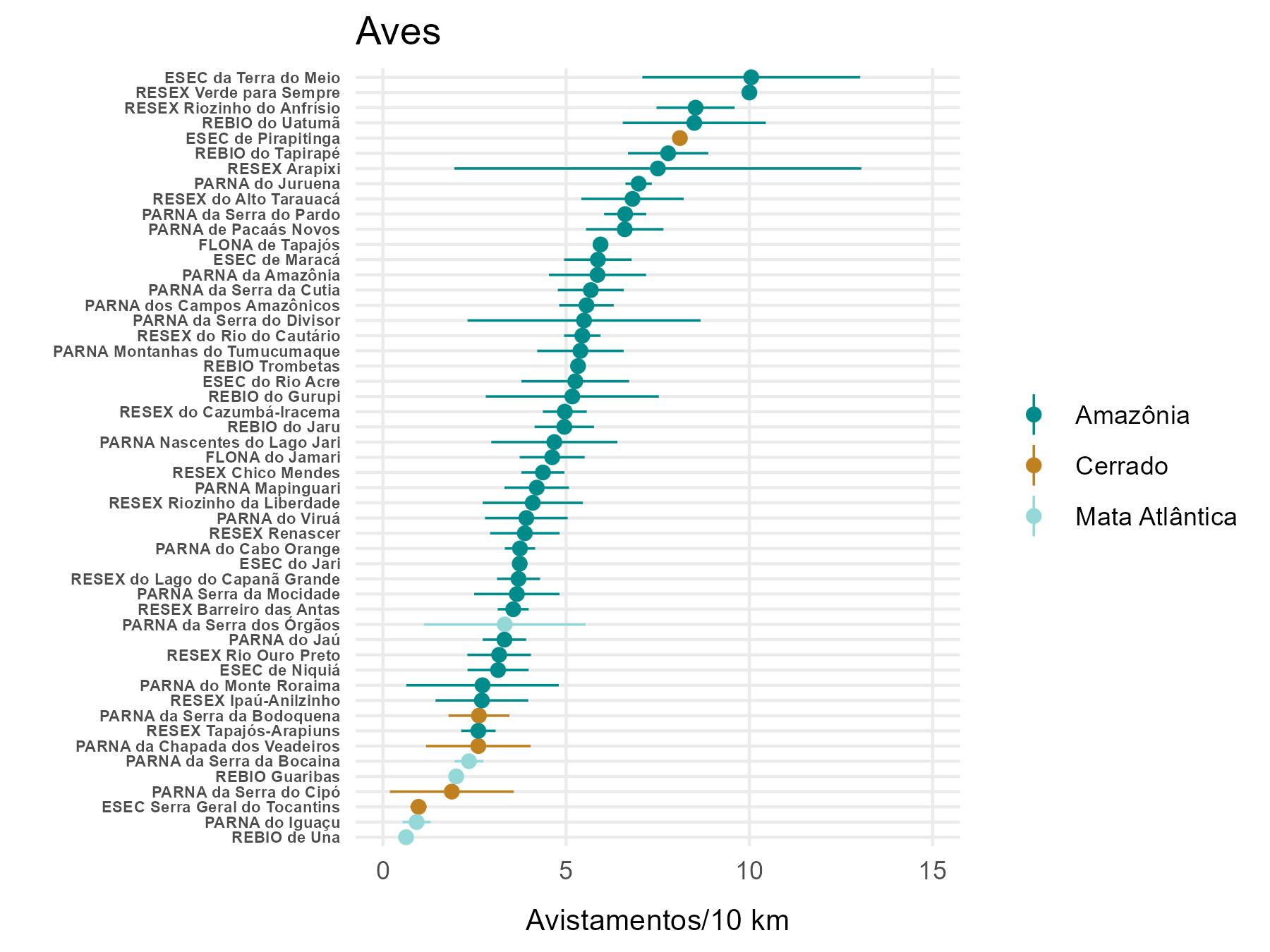
Em termos de abundância total de mamíferos e aves, o bioma Amazônico se destaca por apresentar taxas de avistamento médias mais altas (9,45 e 5,32 avistamentos/10km, respectivamente), com grande variação tanto para mamíferos (1,91 a 20,12) quanto para aves (2,60 a 10,05). No Cerrado as taxas de avistamento variaram de 1,35 a 9,83 e 0,97 a 8,10 para mamíferos e aves, respectivamente (taxas médias de 3,20 e 2,77). Já na Mata atlântica os resultados variaram de 0,63 a 7,50 mamíferos e de 0,63 a 3,32 aves avistadas a cada 10 km, com valores médios de 2,91 e 1,77 (Figura 5.6). Observamos que não foram aplicados testes formais para avaliar a existência de diferenças estatisticamente significativas nas comparações realizadas, sendo apresentada aqui apenas uma análise exploratória e descritiva, com uma interpretação baseada nos valores médios e na inspeção visual dos resultados gerados.
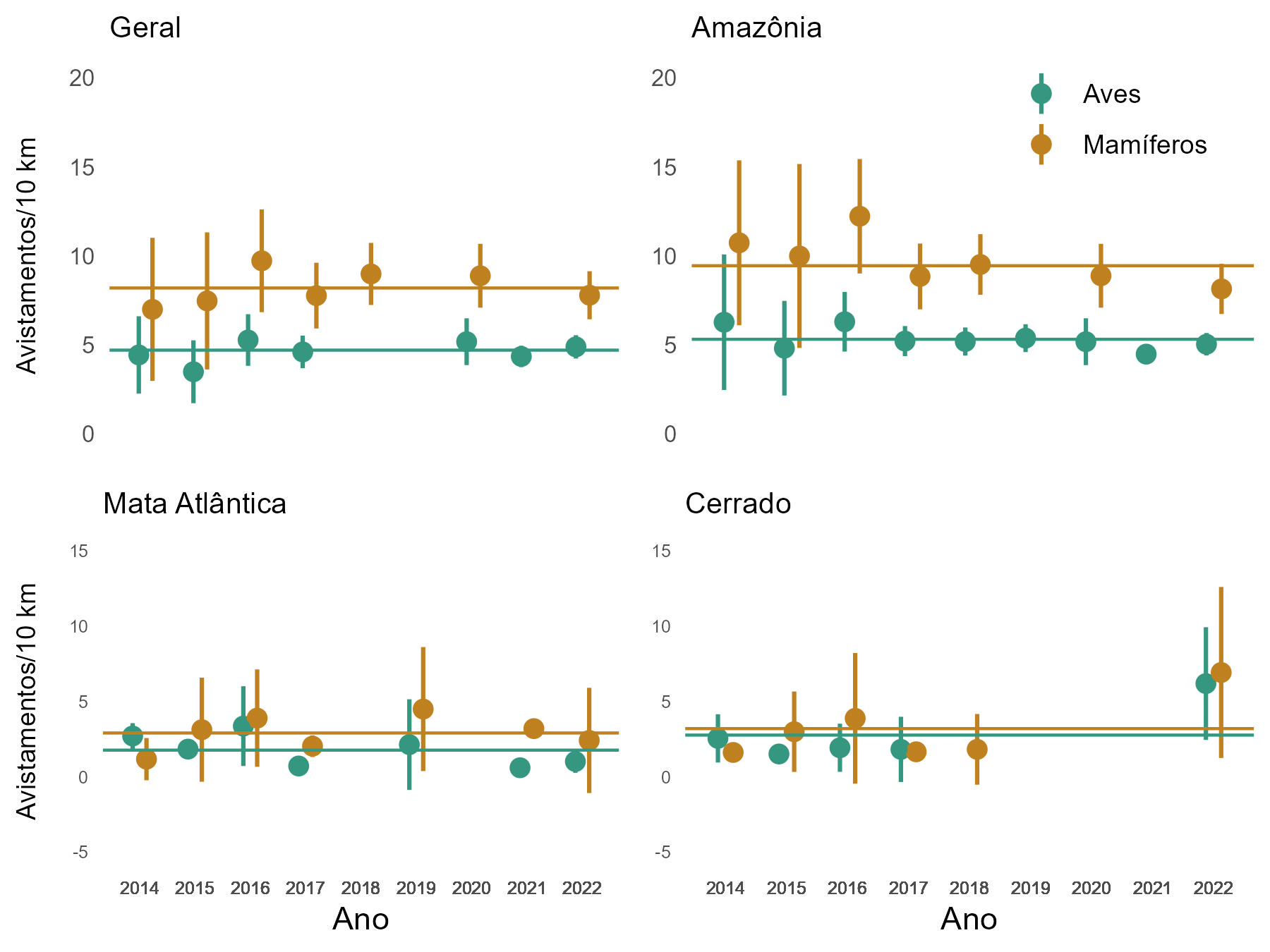
5.2.3 Taxa de encontro de mamíferos e aves ao longo do tempo - geral e por bioma - 2014 a 2022
Considerando a variação da abundância relativa ao longo do tempo, na Amazônia os primeiros anos apresentam uma maior dispersão dos resultados, explicada pelo número proporcionalmente reduzido de UCs participantes e consequente menor esforço amostral na fase inicial do Programa, além de um leve decréscimo nas abundâncias em 2021 e 2022 (Figura 5.7). Para a Mata Atlântica os resultados são irregulares devido à descontinuidade de amostragem e à grande variação no esforço entre os anos. Já para o Cerrado, o padrão observado se justifica pelo fato de duas unidades, o PARNA da Serra da Bodoquena e a ESEC de Pirapitinga, serem distintas das demais, com matriz florestal e, consequentemente, com maior taxa de avistamento.
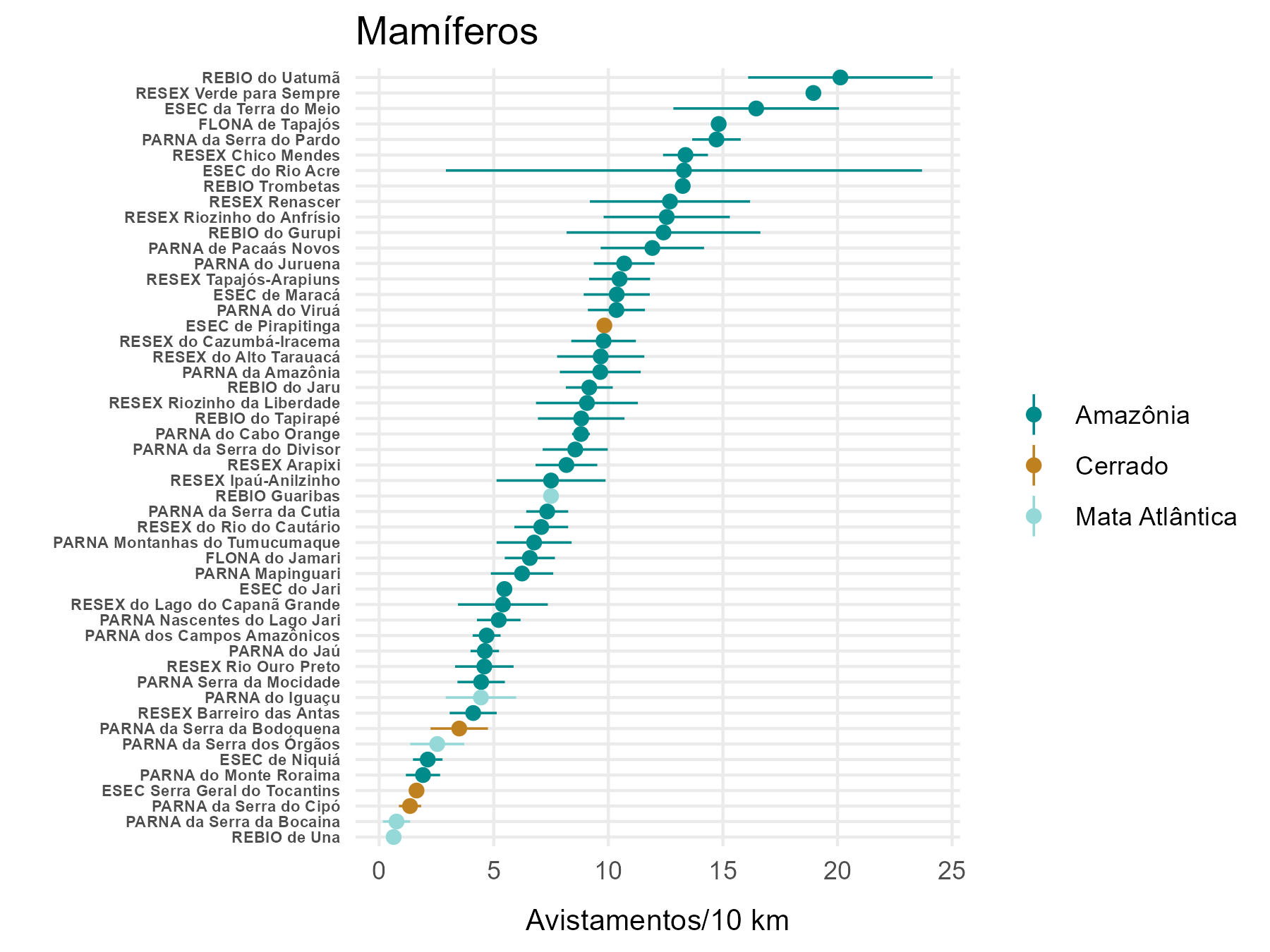
5.2.4 Abundância de mamíferos e aves nas unidades de conservação
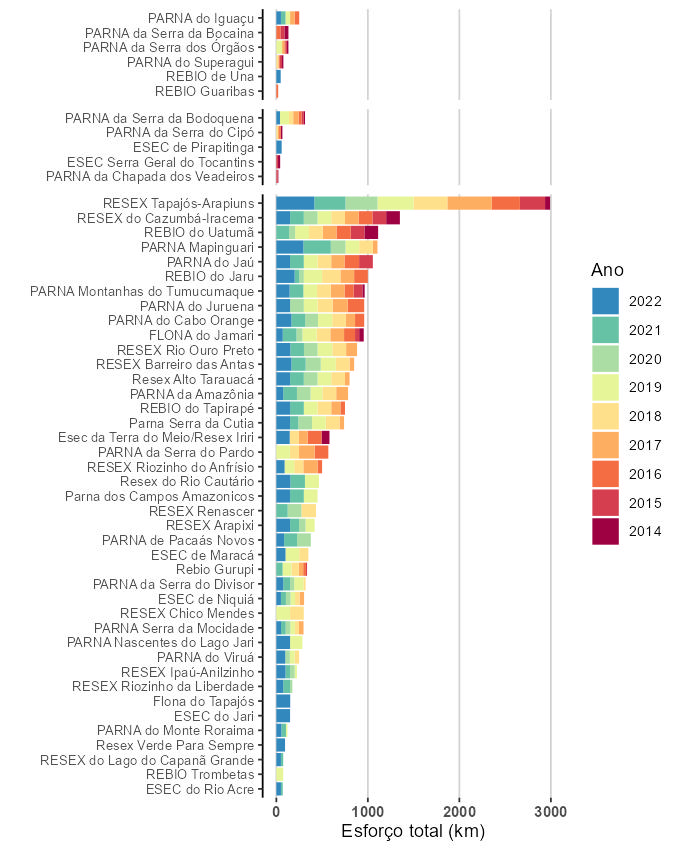
Dentre as unidades de conservação com maiores taxas de avistamento totais para mamíferos destacam-se a REBIO do Uatumã, a RESEX Verde para Sempre e a ESEC da Terra do Meio (Figura 5.8). Essas três UCs também estão entre as quatro com maiores taxas totais de avistamento de aves (Figura 5.9). Contudo, a posição da RESEX Verde para Sempre deve ser considerada com cautela, visto que representa um único ano de amostragem (2022).
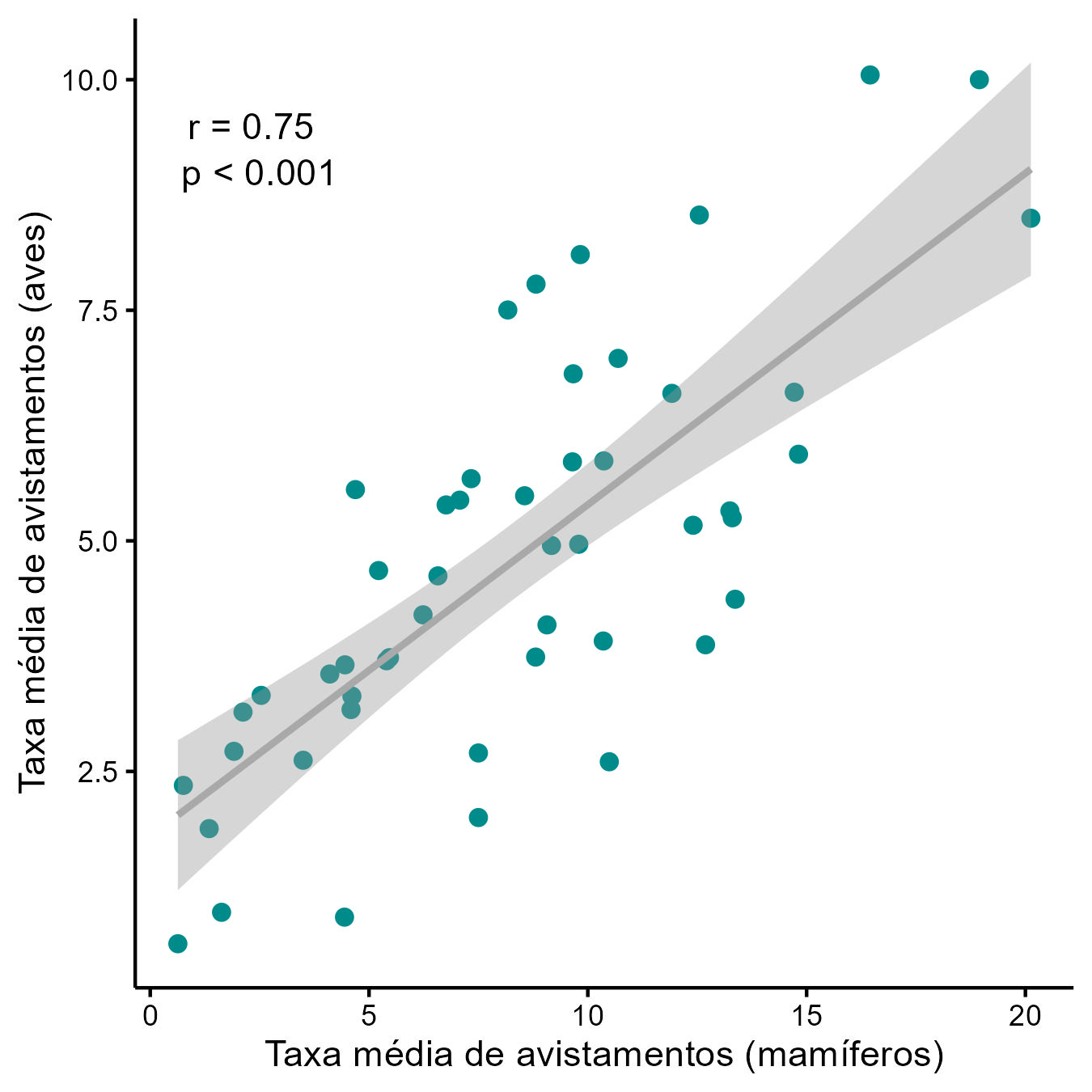
Observou-se uma correlação positiva (r = 0,75; P < 0,001) entre as abundâncias (taxas médias de avistamento) de mamíferos e de aves nas unidades de conservação amostradas (Figura 5.10). Esse resultado não surpreende, visto que o impacto das pressões negativas de origem antrópica deve, em linhas gerais, afetar de forma similar esses dois grupos de vertebrados.
5.2.5 Tendências populacionais de mamíferos e aves em unidades de conservação do Programa Monitora
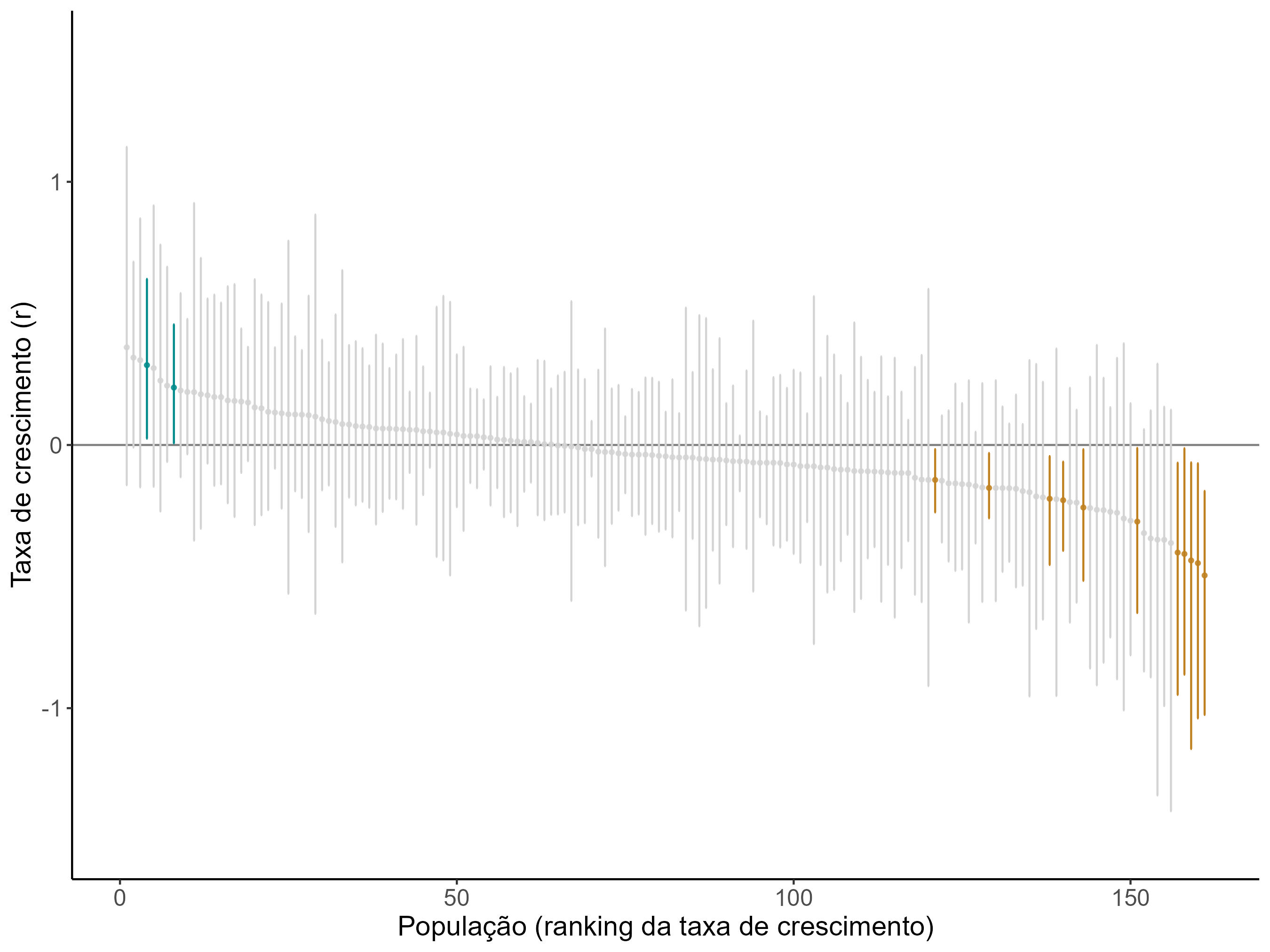
A análise de tendências populacionais foi realizada por meio de modelos de espaço de estados (Auger‐Méthé et al. 2021), tendo como indicador de tendência a taxa de crescimento exponencial r (Daskalova et al. 2020). Basicamente, se r > 0, a população está aumentando, se r = 0, a população está estável, e se r < 0, a população3 está diminuindo.
Para garantir a robustez das análises, somente foram incluídas populações com dados suficientes, com base nos seguintes critérios: (1) série temporal de dados ≥ 5 anos; (2) taxa de encontro média ≥ 0,5 encontros/10 km; (3) esforço amostral anual médio suficiente para atingir coeficiente de variação da taxa de encontro ≥ 0,25.
No total, 89 populações de 36 táxons de mamíferos e 72 populações de 13 táxons de aves, de 21 unidades de conservação, foram selecionadas para análise. A duração do monitoramento para essas populações variou de cinco a nove anos, com média de sete anos. A maioria das populações analisadas (91%, n = 148) permaneceu estável ao longo do monitoramento, com a taxa de crescimento r não diferindo significativamente de zero, enquanto duas populações (1,2%) apresentaram tendência significativa de aumento e 11 (6%) apresentaram tendência significativa de declínio (Figura 5.11, Tabela 5.1).
| Unidade de Conservação | Táxon | Tendência |
|---|---|---|
| Callicebus brunneus | PARNA da Serra da Cutia | ↓ |
| Crypturellus spp. | PARNA da Serra do Divisor | ↓ |
| Dasyprocta fuliginosa | PARNA da Serra do Divisor | ↓ |
| Mico melanurus | PARNA do Juruena | ↓ |
| Tinamus spp. | PARNA do Juruena | ↑ |
| Penelope jacquacu | PARNA do Mapinguari | ↓ |
| Pithecia irrorata | PARNA do Mapinguari | ↓ |
| Myoprocta pratti | RESEX Alto Tarauacá | ↑ |
| Tinamus spp. | RESEX Barreiro das Antas | ↓ |
| Tinamus spp. | RESEX do Cazumbá-Iracema | ↓ |
| Urosciurus spadiceus | RESEX do Cazumbá-Iracema | ↓ |
| Crypturellu spp. | RESEX Rio Ouro Preto | ↓ |
| Sapajus apella | RESEX Rio Ouro Preto | ↓ |
5.2.6 Variação espacial na taxa de avistamento média - mamíferos e aves conjuntamente
O tamanho das unidades de conservação do Programa Monitora varia muito entre biomas. No Cerrado e na Mata Atlântica as unidades de conservação são relativamente pequenas, com algumas dezenas de milhares de hectares, enquanto na Amazônia podem ultrapassar um milhão de hectares. Considerando-se que, nas unidades do Cerrado e Mata Atlântica, a abundância de mamíferos e de aves, expressas pelas taxas de avistamento médias, são inferiores (Figura 5.8 e Figura 5.9), é natural considerar que essas populações apresentem, em geral, uma maior vulnerabilidade demográfica comparada àquela das populações amazônicas. A distribuição espacial da taxa de avistamento média geral (considerando-se conjuntamente mamíferos e aves) é apresentada na Figura 5.12. Dentre as unidades de conservação da Amazônia aquelas localizadas entre os rios Tapajós e Xingu apresentam as maiores abundâncias, à exceção da REBIO do Uatumã, localizada ao norte do rio Amazonas, que também apresenta uma alta taxa média geral de avistamento (Figura 5.12).